
Viajamos al Neolítico para estudiar los cambios en el microbioma que tuvieron lugar, las primeras “domesticaciones” de microbios y cómo el uso de la fermentación marcaría el curso de la historia de la humanidad hasta nuestros días.
La Revolución Neolítica
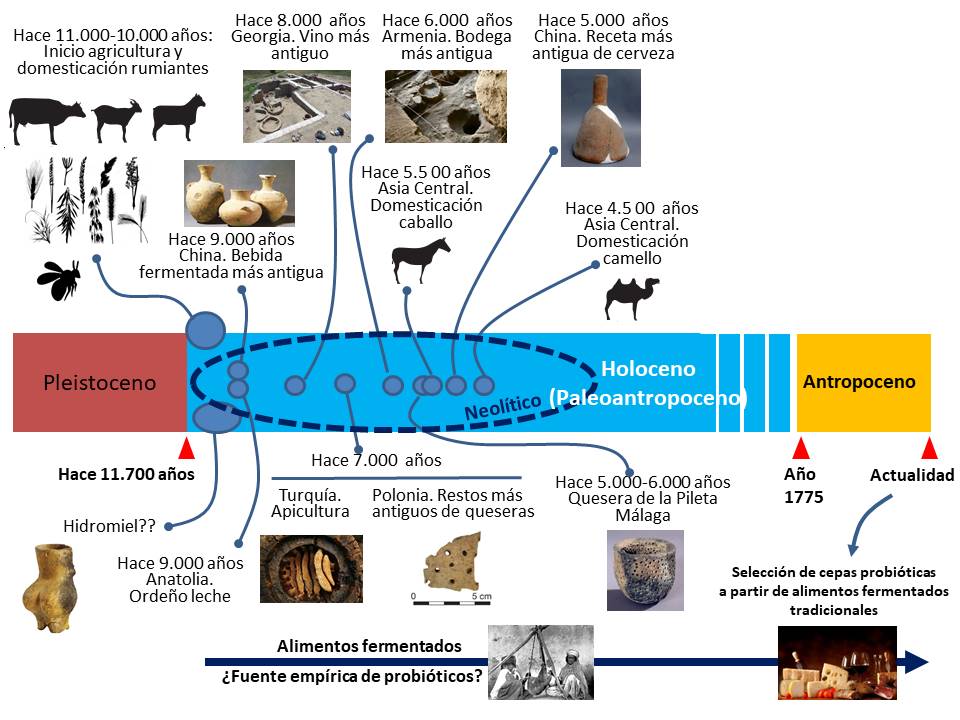
Mucho tiempo antes de que se conociese la existencia de los microorganismos probióticos, la especie humana había empleado numerosos alimentos fermentados, incluyendo el pan, la cerveza, el vino, la sidra, el queso, el kéfir, el kumis, el yogur y otras innumerables variedades de leches fermentadas con fines terapéuticos. En este sentido, la historia de los probióticos se remonta al Neolítico (Ozen y Dinleyici, 2015) (figura 1).
El Neolítico es el periodo de la prehistoria caracterizado por la existencia de útiles de piedra pulimentada, en contraposición a los de piedra tallada propios del Paleolítico, y precede al Calcolítico, que se inicia con la puesta a punto de la tecnología de fundición del cobre. Sin embargo, lo que realmente define a ese periodo es la denominada Revolución Neolítica. Se trata de la primera transformación radical de la forma de vida de buena parte de la humanidad, que pasa de tener una economía cazadora-recolectora a la producción de alimentos a partir de especies vegetales, animales y microbianas domesticadas. Este proceso gradual se inició hace más de 10.000 años y abarcó distintos períodos temporales según cada zona geográfica. Las pruebas más antiguas que atestiguan la transición de unas sociedades de cazadores-recolectores a otras de agricultores y ganaderos se encuentran en Oriente Próximo y datan, aproximadamente, del año 8.500 a.C. El Neolítico en la Península Ibérica se desarrolló, aproximadamente, entre el 6.000 a.C. y el 3.000 a.C. (Szécsényi-Nagy et al., 2017).
Con esta nueva economía, la población comienza a establecerse permanentemente en un lugar: aparece el sedentarismo. Las tareas agrícolas y ganaderas propiciaron el desarrollo de nuevos útiles (azadas, hoces, molinos de mano…), de tecnologías (roturación de la tierra, almacenamiento de cosechas, manejo de los animales) y de artesanías como la cestería y, sobre todo, la cerámica, que fue primordial para el tratamiento y la conservación de los alimentos. Los cambios ligados a la Revolución Neolítica significaron un enorme salto para el progreso de la humanidad, que comenzó a crecer con mayor rapidez al acumular alimentos que podían conservarse durante bastante tiempo. La aparición de excedentes permitió la especialización y división del trabajo, la acentuación de las diferencias sociales y la aparición del comercio.
El Neolítico también conllevó una alteración progresiva pero radical del medio ambiente, que se convierte en un paisaje humanizado de tipo agrícola, ganadero y forestal. La intervención humana en el desarrollo y propagación de las especies vegetales y animales domesticadas permitió una prolongada selección artificial, eminentemente empírica, que dio lugar a un enorme incremento de la diversidad genética (variedades, razas…).
Cambios en el microbioma asociados al Neolítico
Desde entonces, la influencia humana sobre la atmósfera, la hidrosfera y la biosfera del planeta ha sido de tal magnitud que, de hecho, justifica una nueva época geológica: el Antropoceno. Estos cambios también afectaron a nuestra microbiota. Los recientes avances en ecología y genómica microbiana muestran que las grandes perturbaciones de las poblaciones microbianas humanas se corresponden estrechamente con las grandes fases del Antropoceno: a) el “Paleoantropoceno”, que comenzó precisamente con el surgimiento de la agricultura y la ganadería; b) la Revolución Industrial, a finales del siglo XVIII; y c) la "gran aceleración", desde la década de 1950 hasta la actualidad (Guillins y Paulsen, 2014).
La domesticación de ciertas plantas y animales, además de garantizar un suministro más continuo y estable de alimentos, redujo la diversidad de la dieta neolítica respecto a la paleolítica. El análisis de la microbiota oral a partir de restos dentales fósiles ha mostrado un marcado cambio en la diversidad y composición bacteriana durante la transición de un estilo de vida cazador-recolector hacia un mayor consumo de cereales durante el Neolítico (8.000 años a.C.) (Adler et al., 2013). Por otra parte, el análisis del microbioma conservado en diversos coprolitos (heces fósiles) ha confirmado que la dieta afectó drásticamente a la composición del microbioma y sugiere que los microbiomas “neolíticos” eran similares a los de las comunidades rurales modernas, que conservan dietas ricas en fibra y polisacáridos (De Filippo et al., 2010; Walter y Ley, 2011; Yatsunenko et al., 2012).
En el pasado, se creía que los microorganismos eran cosmopolitas, porque su tamaño pequeño, sus grandes poblaciones y su capacidad para permanecer en un estado “durmiente” favorecían su fácil dispersión. Actualmente se considera que la difusión espacial de muchos microorganismos sigue ciertos patrones, similares a los exhibidos por los macroorganismos (Green et al., 2004; Horner-Devine et al., 2004). En este sentido, las actividades humanas influyen notablemente en la dispersión y concentración de virus, arqueas, bacterias, hongos, protozoos y helmintos, independientemente de que actúen como patógenos obligados u oportunistas, comensales o mutualistas (Litchman, 2010; Wilkinson, 2010). El primer transporte de microorganismos ligado al hombre ocurrió cuando nuestra especie se dispersó desde África, llevando consigo sus microorganismos y, por supuesto, sus parásitos (Michelet y Dauga, 2012). De hecho, el tráfico de parásitos humanos se está utilizando para rastrear las migraciones humanas durante la Prehistoria (Araujo et al., 2008).
La domesticación de los animales y el desarrollo de la agricultura, además de modificar la dieta, también provocaron otros grandes cambios en la ecología microbiana humana. Los asentamientos agrícolas y el aumento consiguiente de la densidad de la población facilitaron el contagio de las enfermedades infecciosas y lo hicieron en un ambiente donde el contacto con animales era cotidiano. Esto permitió que diversos patógenos del ganado se adaptaran e infectaran a los huéspedes humanos (y viceversa), aumentando drásticamente el impacto de las zoonosis en una población humana crecientemente sedentaria. Ciertas enfermedades víricas (como las paperas, el sarampión, la gripe o la viruela), bacterianas (como la difteria y la tos ferina) o parasitarias (malaria causada por Plasmodium falciparum) parecen haberse originado de este modo. Por otra parte, los asentamientos permanentes también crearon nuevos hábitats para vectores de enfermedades, incluyendo ciertas especies de roedores, aves y artrópodos (insectos y arácnidos hematófagos) (Smith y Guégan, 2010). En consecuencia, estos factores contribuyeron al cambio de la microbiota humana y a la “endemización” de algunas enfermedades. Por otra parte, la pérdida de diversidad vegetal inherente a la agricultura facilitó la expansión de plantas invasoras ("malas hierbas") y, con ello, la de microorganismos asociados a todas ellas, tanto beneficiosos (micorrizas y rizobios) como fitopatógenos, un fenómeno que continúa en nuestros días (Pringle et al., 2009; Rodríguez-Echeverría, 2010).
Alimentos fermentados: la “domesticación” de los microbios
Como ya se ha señalado, la Revolución Neolítica supuso un aumento de la disponibilidad de alimentos y, como consecuencia, la aparición y desarrollo de diversas técnicas para el tratamiento y la conservación de dicho excedente (cocción, secado, salazón, ahumado…). Entre esas técnicas (que se pueden combinar entre sí), la fermentación ha ocupado un lugar destacado desde entonces hasta nuestros días, haciendo “de la necesidad virtud”.
Los primeros agricultores y ganaderos observaron que si almacenaban en recipientes impermeables alimentos azucarados como las frutas, los granos o la leche, podían suceder dos cosas: en ocasiones se generaban productos de mal olor y sabor cuyo consumo resultaba tóxico (lo que denominamos hoy putrefacción), pero en otras se producían derivados de sabor acre pero no desagradable, que no afectaban a la salud y que fueron los precursores de la sidra, el vino, la cerveza o el queso. Aunque no sabían el porqué de estos procesos, sobre la base de un sistema empírico de prueba y error, y con una gran capacidad de observación, poco a poco fueron controlando las condiciones de tiempo, temperatura, atmósfera, etc. que evitaban la putrefacción y conducían hacia lo que más tarde se denominó “fermentación”. La palabra “fermentación” procede del latín “fervere”, que significa “hervir”. El nombre proviene de la observación de que los mostos procedentes del pisado de las uvas producían burbujas, como si estuvieran hirviendo.
Los alimentos fermentados debieron ser rápidamente apreciados, más allá de la (ya de por sí) trascendental transformación de materias primas muy perecederas en productos con una vida útil mucho más prolongada. Las fermentaciones proporcionaban nuevos alimentos con una amplia gama de sabores, olores y texturas, y en muchos casos, más digeribles que las materias de partida. Algunos de ellos contenían cantidades importantes de una sustancia tan poderosa que realmente ejercía efectos “mágicos” (etanol). Pero, además (y a tenor de lo observado en pueblos con una forma de vida similar a la del neolítico), algunos de los productos fermentados se debieron emplear con fines medicinales por dos razones principales: eran muy nutritivos y, aunque entonces no se sabía, contenían grandes cantidades de microbios con propiedades beneficiosas para la salud (Nakazawa y Hosono, 1992).
Una práctica que se reveló muy útil para promover la fermentación y evitar la putrefacción fue la adición de una porción de producto con características deseables a la materia prima que se deseaba fermentar (lo que tradicionalmente denominamos “fermento” o “masa madre”). Esta práctica provocó la selección de comunidades microbianas adaptadas a las características del alimento a fermentar, provocando una rápida especialización genómica a través de mecanismos tales como la pseudogenización, la eliminación, modificación o duplicación de ciertos genes, la hibridación interespecífica o la transferencia horizontal de genes (Sicard y Legras, 2011; Gibbonsy Rinker, 2015; Douglas y Klaenhammer, 2010). Dentro de los nuevos nichos alimentarios, los requerimientos metabólicos de esos microbios se volvieron predecibles, resultando en la “domesticación” de comunidades microbianas con capacidades fermentativas cada vez mejores, que se desarrollaban en la masa fermentable y producían una serie de compuestos que, por un lado, impedían la colonización por organismos alterantes o patógenos y que, por otro, daban lugar a efectos beneficiosos para las personas que los ingiriesen. Asistimos, pues, al nacimiento de la fabricación de productos biotecnológicos funcionales con propiedades probióticas.
Evidencias arqueológicas más antiguas de fermentaciones alimentarias
Hasta hace pocos años se pensaba que las primeras fermentaciones dirigidas por la especie humana se produjeron hacia el año 2.000 a.C. Sin embargo, la aplicación de las modernas técnicas químicas y de biología molecular a la arqueología ha mostrado que ya se realizaban, de forma sistemática, varios milenios antes (figura 1).
El análisis químico de compuestos orgánicos absorbidos en vasijas de cerámica de la antigua aldea neolítica de Jiahu (China) ha revelado que en el séptimo milenio a.C. ya se producía una bebida fermentada a partir de una mezcla de arroz, miel y fruta (majuelos y/o uvas silvestres) (McGovern et al., 2004) (figura 2). El papel de cereales, frutas, leche e incluso carnes en las fermentaciones alimentarias primitivas es bien conocido. Sin embargo, el de la miel lo es mucho menos, cuando no ignorado. Este alimento fue muy apreciado durante milenios (a menudo reservado para las élites) y tenía un extenso uso con fines religiosos y medicamentosos (parte de los cuales podían deberse a su gran contenido en sustancias potencialmente prebióticas). Las abejas y los humanos han disfrutado de una larga asociación, como lo demuestra la iconografía en el arte rupestre (figura 3). Recientemente se ha observado un uso continuo y ampliamente extendido de la miel en Europa, Oriente Medio y África del Norte durante más de 8.000 años, por lo que todo parece indicar que las abejas se domesticaron al mismo tiempo que los mamíferos (Roffet-Salque et al., 2015). La vasija más antigua con restos de cera de abejas se encontró en Çatalhöyük (Turquía) y tenía casi 9.000 años de antigüedad. Actualmente se considera que la hidromiel o aguamiel (bebida que se obtiene a partir de la fermentación de una mezcla de agua y miel) fue la primera bebida alcohólica que consumió el hombre, y se cree que es precursora del vino y la cerveza (Toussaint-Samat, 2008). En este contexto, resulta interesante señalar que diversas especies de lactobacilos y de bifidobacterias osmotolerantes y resistentes a los componentes antimicrobianos del polen y de la miel se encuentran entre los microorganismos dominantes en la microbiota del polen, del aparato digestivo de las abejas y de la miel (Corby-Harris et al., 2014; Alberoni et al., 2016; Kwong y Moran, 2016). Recientemente se ha descrito que la adición de miel al rayeb (una leche fermentada tradicional de Oriente Medio) aumenta significativamente la viabilidad de lactobacilos y bifidobacterias (Ismail et al., 2017).
El vino (alimento, medicina, droga, vínculo social, símbolo religioso y de poder…) está íntimamente asociado a la civilización tal como la conocemos en Occidente. Por lo tanto, no es de extrañar que el interés por conocer su origen haya sido mayor que el del hidromiel. Recientemente se han encontrado residuos de vino en fragmentos de cerámica procedentes de dos yacimientos arqueológicos de Georgia (zona al sur del Cáucaso) que datan de aproximadamente el año 6.000 a.C. (McGovern et al., 2017) (figura 4). El equipo detectó la presencia de los ácidos tartárico, málico, succínico y cítrico (combinación que únicamente se encuentra en el vino), siendo la evidencia más temprana hasta la fecha de elaboración de dicha bebida (aproximadamente 8.000 años de antigüedad). Se descubrió así que el vino se fermentaba y envejecía en grandes vasijas (de unos 300 litros), que se enterraban hasta el cuello para conseguir la estabilización de la temperatura de fermentación, práctica que sigue siendo habitual en Georgia en la actualidad (vino quevri). Previamente, el equipo de McGovern ya había encontrado pruebas químicas de producción vinícola, de aproximadamente 7.000 años de antigüedad, en una cueva de Godin Tepe (Montañas Zagros, Irán) y los restos de una auténtica bodega (lagar, vasijas, copas, restos de uvas) en una cueva de Areni, en la actual Armenia (aproximadamente 6.000 años de antigüedad) (figura 5). Curiosamente, se trata de la misma cueva donde se encontró el zapato más antiguo conocido, un mocasín de cuero bien conservado de 5.500 años de antigüedad. Areni es un pueblo armenio remoto, próximo al Monte Ararat, donde se dice que Noé plantó un viñedo, cosechó uvas, las fermentó y se emborrachó.
La cerveza fue (y sigue siendo) una bebida fermentada muy consumida y, al igual que el vino, parece tener su origen en el Neolítico. Una investigación reciente ha revelado la existencia de una receta para fabricar cerveza de mijo de 5.000 años de antigüedad en Yangshao (la región conocida como “la cuna de la civilización china”). A este cereal se le añadía, además, cebada, lágrimas de Job y tubérculos. Los elaboradores usaban herramientas especializadas para machacar los granos, llevaban a cabo el malteado y controlaban la temperatura del proceso, demostrando que ya por entonces existía una técnica muy avanzada de elaboración de esta bebida (Wang et al., 2016) (figura 6). La adición de cebada al mijo se hacía, presumiblemente, para incrementar la concentración de α y β-amilasa, enzimas que promueven el proceso de sacarificación (conversión del almidón en azúcar), mientras que los tubérculos aportarían más almidón y azúcares para la fermentación y conferirían un sabor más dulce a la cerveza. El cuarto milenio antes de Cristo se caracterizó en aquella zona por una creciente organización jerárquica, en la que eran frecuentes los festines rituales organizados por las élites y que implicaban un consumo abundante de alcohol. Distintos autores han sugerido que la producción y el consumo de vino, cerveza y otras bebidas fermentadas alcohólicas pueden haber contribuido al surgimiento de sociedades jerárquicas sobre una base político-religiosa.
Leche y productos lácteos: un salto cualitativo para la humanidad
Si hay un tipo de alimento que se haya asociado por excelencia con la presencia de microorganismos potencialmente probióticos, ése son las leches fermentadas (Mayo et al., 2010). La domesticación de los animales fue simultánea a la de los vegetales; probablemente la única excepción sería la del perro, que parece ser muy anterior (hace entre 20.000 y 40.000 años), una especie propia de sociedades cazadoras (Botigué et al., 2017). Obviamente, el ganado ovino, caprino y vacuno (originario de Oriente Próximo y Medio) y el equino (estepas de Asia Central) fueron los más importantes por lo que respecta a su producción láctea (aparte de los restantes múltiples usos que todos ellos tuvieron); actualmente no existen pruebas de un uso antiguo de los camélidos domésticos (dromedarios, llamas y alpacas) para tal fin, a excepción de la leche de camella en las estepas de Mongolia (donde se dejaba y se sigue dejando fermentar para producir el khoormog).
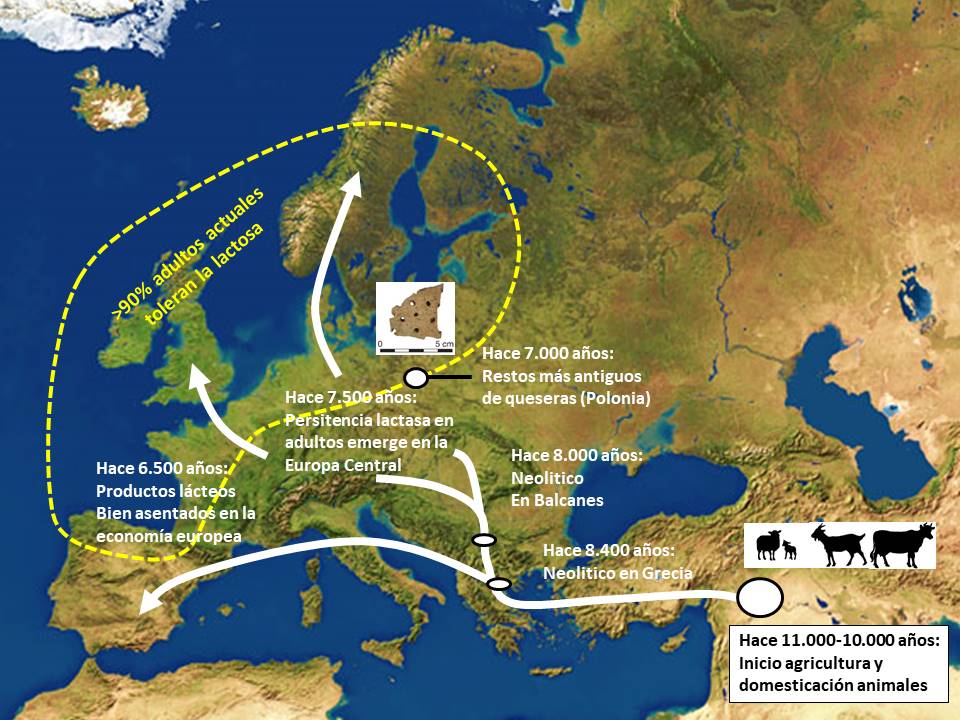
La domesticación del ganado bovino, ovino y caprino tuvo lugar en el Cercano Oriente durante en el octavo milenio a.C. (Zeder et al. 2006). El análisis de los residuos orgánicos conservados en fragmentos de 2.200 vasijas procedentes de numerosos yacimientos arqueológicos del Cercano Oriente y del sureste de Europa ha revelado que ya se ordeñaba en el séptimo milenio a.C., siendo esta actividad particularmente importante en el noroeste de Anatolia (Evershed et al., 2008). Otros autores han mostrado evidencias del uso de leche de rumiantes domésticos en el sexto milenio a.C. en Europa oriental (Craig et al., 2005), en el quinto milenio a.C. en el norte de África (Dunne et al., 2012) y en el cuarto milenio a.C. en la actual Gran Bretaña (Copley et al., 2003). En contraste con el Neolítico de Eurasia, el norteafricano (en un entonces “Sahara verde”) se caracterizó por una dependencia del ganado bovino, ovino y caprino como una forma de vida estable y extendida, mucho antes de la primera evidencia de plantas domesticadas o de comunidades establecidas en aldeas (Marshall y Hildebrand, 2002; Gifford-Gonzalez y Hanotte, 2011).
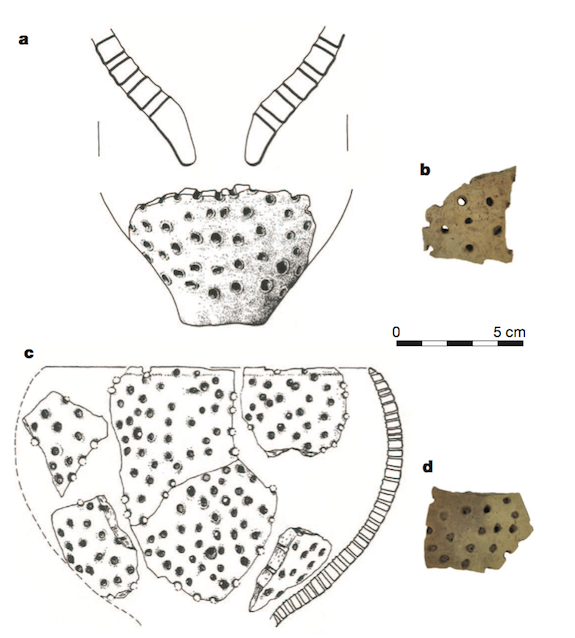
Sin embargo, una cosa es encontrar restos de leche y otra distinta tener evidencias de la fabricación de productos lácteos fermentados. El enlace entre ambos acontecimientos se produjo en los años 20 y 30 del siglo XX, cuando los arqueólogos comenzaron a descubrir extrañas piezas de cerámica llenas de agujeros en yacimientos neolíticos de Polonia de unos 7.000 años de antigüedad, y cuya utilidad era completamente desconocida. En la década de 1970, el arqueólogo Peter Bogucki estudió ese tipo de fragmentos e inmediatamente le vinieron a la mente los coladores (encellas) que había visto en casa de un amigo quesero, por lo que relacionó esos artilugios con la fabricación de queso (figura 7). Desafortunadamente, no tenía manera de probar su idea en aquel momento. Los misteriosos fragmentos de cerámica quedaron almacenados hasta 2011, cuando el equipo de Richard Evershed los analizó, encontrando numerosos restos de grasas lácteas (Salque et al., 2013); en otras palabras, los primeros agricultores habían utilizado esos objetos como coladores para separar la cuajada del suero líquido, convirtiendo a las reliquias polacas en la evidencia más antigua conocida de la fabricación de queso en el mundo (figura 8).
El “procesado” de la leche, particularmente la producción de quesos, mantequilla y leches fermentadas, supuso un hito para la humanidad porque no solo permitió su conservación de forma no perecedera y transportable, sino que también la convirtió en un producto más digerible para los primeros ganaderos prehistóricos. Por entonces, los adultos no podían tomar leche porque, a diferencia de los niños lactantes, no disponían de la enzima lactasa necesaria para descomponer la lactosa, su principal azúcar (Burger et al., 2007). La fermentación lleva implícita la conversión de la lactosa en ácido láctico, por lo cual la intolerancia asociada a su consumo disminuye, haciendo fácilmente digerible el producto resultante (Curry, 2013).
Siglos después, algún habitante de las amplias llanuras de la actual Hungría sufrió una mutación genética que le proporcionó una gran ventaja respecto a sus congéneres y que tendría importantes consecuencias para sus descendientes. En el ADN de esta persona se produjo un cambio de un nucleótido por otro (citidina a timidina) en las proximidades del gen que codifica para la lactasa, que así paso a producirse constitutivamente (es decir, durante toda la vida del individuo y no solo durante los dos-tres primeros años de vida) (alelo LP o “Lactose Persistence”) (Itan et al., 2009). Esto le permitió tener acceso a un alimento prácticamente completo y fácil de obtener en una sociedad de pastores, por lo cual dicha mutación dio una importante ventaja a los individuos que la portaban y se diseminó muy rápidamente por toda Europa, encontrándose ya, hace 6.500 años, en el norte de Alemania. De hecho, el grado de selección fue tal que se considera entre los más potentes observados hasta la fecha para cualquier gen humano (Bersaglieri et al., 2004). De este modo, a principios del periodo Calcolítico, hace unos 5.000 años, el alelo LP era prevalente en la mayor parte de Europa, y el pastoreo del ganado se había convertido en una parte dominante de la cultura. Hoy en día, la mayoría de las personas que conservan la capacidad de digerir la leche en la edad adulta tienen ascendencia europea.
Durante mucho tiempo, la corriente principal de la arqueología europea pensaba que los cazadores-recolectores mesolíticos europeos se convirtieron gradualmente en agricultores neolíticos. Sin embargo, las conclusiones del proyecto multidisciplinario “LeCHE” (Lactase Persistence in the Early Cultural History of Europe), que incluyó análisis paleogenómicos humanos y análisis de huesos de animales domésticos hallados en distintos yacimientos, son completamente diferentes. Sus resultados sugieren que los agricultores-ganaderos de Oriente Medio y Próximo se expandieron rápidamente por Europa gracias a su tecnología, desplazando a la población cazadora-recolectora local (Bramanti et al., 2009; Ethier et al., 2017); además, en lugar de gastar esfuerzos en domesticar a los rumiantes autóctonos (como el uro), trajeron su ganado con ellos (Edwards et al., 2007). En otras palabras, los nuevos colonos agrícolas superaron a los lugareños gracias a una combinación de genes y tecnología agropecuaria y alimentaria que les proporcionaba una mayor vitalidad, una mayor tasa de fertilidad y una mayor esperanza de vida. Y a medida que los sureños avanzaban hacia el norte, el alelo LP “surfeaba” en la ola de migración (Curry, 2013) (figura 9).
Algunos de los participantes del consorcio LeCHE actualmente forman parte del proyecto BEAN (Bridging the European and Anatolian Neolithic), que analiza quiénes fueron los primeros agricultores y pastores y cuándo y cómo llegaron a Europa. En su trabajo de campo en Anatolia se encontraron con el beyazpeynir, un queso salado de leche de oveja, cabra o vaca que se come en casi todos los desayunos de la zona. Probablemente es muy parecido al queso que los granjeros neolíticos de la región habrían comido hace unos 8.000 años, mucho antes de que la persistencia de la lactasa permitiera a las personas beber leche fresca.
El proyecto LeCHE constituye un modelo de investigación que podría aplicarse a otros cambios muy relevantes en la dieta humana, incluyendo nuestra adaptación a otros muchos alimentos fermentados. Un enfoque similar podría, por ejemplo, ayudar a desvelar los orígenes de la amilasa, una enzima que ayuda a descomponer el almidón y que podría haber sido causa o consecuencia del creciente apetito por el grano que acompañó al desarrollo de la agricultura. Igualmente se podría rastrear la evolución de la alcohol-deshidrogenasa, que es crucial para la descomposición del etanol y podría revelar los orígenes y difusión de la gran afición de nuestra especie por las bebidas alcohólicas.
La importancia de la leche de yegua en las leches fermentadas del Asia Central
La domesticación del caballo se produjo hace aproximadamente 5.500 años en el Kazajstán actual (Librado et al., 2016), a una considerable distancia espacial y temporal de los centros de domesticación de rumiantes (Zeder et al. 2006). Los caballos fueron utilizados inicialmente como fuente de carne y leche (Outram et al., 2009) pero, a diferencia de los rumiantes, también revolucionaron la guerra, el comercio y el intercambio de mercancías, personas e ideas, teniendo una influencia decisiva en el devenir sociopolítico y económico de las sociedades humanas (Kelekna, 2009).
Desde la domesticación equina hasta nuestros días, la leche de yegua ha sido la materia prima esencial para la elaboración de numerosas variedades de leches fermentadas en las extensísimas llanuras y estepas de Asia Central, desde las estribaciones orientales de los Urales hasta China, pasando por Mongolia. En general, todos esos productos lácteos se suelen conocer hoy día bajo la denominación genérica de kumis (también conocidos como koumiss, kumys, kymys o tarag) (figura 10). Se trata de un tipo de alimentos muy rico en nutrientes y que parece haber sido esencial para la supervivencia y estabilidad de las comunidades humanas en unas condiciones climatológicas muy adversas. Las poblaciones locales siempre consideraron que su consumo tenía propiedades medicinales y posiblemente, fue el primer tipo de leches fermentadas cuyas propiedades beneficiosas para la salud fueron recogidas en tratados y revistas médicas con prestigio internacional. El primer artículo al respecto se publicó en el siglo XVIII (Grieve, 1788) y fue seguido de una serie de tres artículos publicados entre 1872 y 1877 en el British Medical Journal (Jagielski, 1872; Jagielski, 1874; Jagielski, 1877) y otro en 1875 en el entonces denominado Western Lancet (figura 11).
Sin embargo, la popularidad que alcanzarían el kéfir y, especialmente, el yogur en los siguientes años (productos cuya historia se abordará en un capítulo independiente) eclipsó al kumis, un producto que ha permanecido prácticamente invisible a los ojos de la ciencia e industria alimentaria occidental. La utilización de los mismos odres durante décadas e incluso siglos, contribuye a la formación de biopelículas espesas y complejas en sus superficies internas, dominadas por bacterias lácticas y levaduras. En las últimas décadas, diversos trabajos científicos han mostrado la extraordinaria diversidad y abundancia de este tipo de microbios en el kumis (Gesudu et al., 2016; Yao et al., 2017) y han confirmado que su consumo regular puede modular la microbiota (Zhang et al.; 2014a,b) y proporcionar ciertos beneficios para la salud (Tsenina y Frolov, 1980; Bakir et al., 2015); además, se han caracterizado las propiedades probióticas deciertas cepas aisladas a partir de kumis tradicional (Wu et al., 2009; Wu et al., 2010; Zhang et al., 2010; Pan et al., 2011; Zhao et al., 2014; Rong et al., 2015) y algunas de ellas se han seleccionado para la elaboración de productos lácteos más modernos o para su venta como nutracéuticos.
Alimentos fermentados tradicionales y probióticos
Los alimentos fermentados tradicionales son el hogar de consorcios microbianos más o menos complejos, cuyos componentes se han seleccionado de una forma empírica durante milenios a partir de la microbiota indígena natural de las materias primas vegetales o animales de partida, de los utensilios y recipientes empleados para su elaboración y/o del medio ambiente en el que se producen. Globalmente, la diversidad de los microorganismos asociados a estos procesos es enorme e incluye mohos, levaduras, bacterias e incluso arqueas. Estos consorcios modifican la composición química de las materias primas de partida para convertirlas en alimentos más estables y apetecibles y, en muchos casos, en verdaderos alimentos funcionales, con mayor valor nutricional y otros beneficios para la salud de los consumidores (Campbell-Platt, 1994; Steinkraus, 1997; Tamang, 2010b; Tamang et al., 2016a,b).
Por otra parte, la diversidad genética asociada a la ingente cantidad de cepas distintas que se pueden aislar a partir de estos alimentos, es tan extraordinaria como desconocida, lo mismo que sucede con su potencial biotecnológico o probiótico. En los últimos años algunos grupos de investigación están haciendo un esfuerzo para el aislamiento y la caracterización de las propiedades probióticas de cepas aisladas de alimentos fermentados tradicionales que nunca se habían estudiado (o muy escasamente) hasta la fecha (Dewan yTamang, 2007; Tamang et al., 2009; Shi et al., 2012; Franz et al., 2014; Kumari et al., 2016). En este sentido, los alimentos fermentados, junto con las muestras biológicas humanas (heces, exudado vaginal, cavidad oral, leche…), han sido las dos grandes fuentes de los microorganismos probióticos comercializados en la actualidad.
La fermentación tradicional de los alimentos representa un patrimonio cultural extremadamente valioso (Tamang et al., 2016a). Cada comunidad en el mundo tiene una cultura alimentaria distinta, que incluye alimentos fermentados y bebidas alcohólicas que suelen simbolizar el patrimonio y los aspectos socioculturales de cada etnia y/o región (figura 12). Sin embargo, numerosos alimentos fermentados tradicionales han desaparecido o su consumo ha disminuido drásticamente en las últimas décadas. Por una parte, los cambios generalizados en los estilos de vida han afectado notablemente a los hábitos alimentarios y a las prácticas culinarias tradicionales, especialmente en los países occidentales. La globalización ha conllevado un consumo mucho mayor de alimentos procesados y el empleo de cultivos iniciadores comerciales para la estandarización de los procesos fermentativos. Por otra parte, los cambios climáticos y medioambientales producidos en los últimos siglos o décadas en algunas zonas del planeta, como la región del Sahel en África o las vastas áreas adyacentes al desierto de Gobi en Asia, también han contribuido a una reducción del espectro de alimentos fermentados tradicionales.
A pesar de ello, todavía existen unos 3.500 alimentos fermentados (incluyendo las bebidas) en el mundo, cifra que puede aumentar hasta unos 5.000 cuando se incluyen variedades con una difusión muy reducida (Campbell-Platt, 1987; Tamang, 2010b; Bourdichon et al., 2012;). Globalmente, los productos fermentados son consumidos hoy en día por miles de millones de personas, ya sea como alimentos básicos o como complementos de la dieta (Tamang, 2010b). Estos alimentos se han clasificado en nueve grandes grupos según las materias primas utilizadas y/o la forma de producción: 1) cereales; 2) vegetales verdes y brotes de bambú; 3) leguminosas; 4) tubérculos y raíces; 5) productos lácteos; 6) productos cárnicos; 7) productos de la pesca; 8) otros alimentos; y 9) bebidas alcohólicas (Steinkraus, 1997; Tamang, 2010b,c). En la tabla 1 se recoge una lista con algunos de los principales alimentos fermentados que actualmente se siguen produciendo de forma artesanal en el mundo. Muchos de ellos son similares a aquellos que se empezaron a elaborar durante el Neolítico y que gracias a sus propiedades nutricionales, funcionales y organolépticas, y a su arraigo en las sociedades en las que se producen han llegado con plena vigencia hasta nuestros días, tanto en los países en vías de desarrollo como en los desarrollados. Todos ellos son fuentes atractivas para el aislamiento de cepas probióticas.
Agradecimientos
El autor desea mostrar su mayor agradecimiento al Dr. Juan Evaristo Suárez (Universidad de Oviedo) por su excelente revisión del capítulo.
Referencias bibliográficas
- Adler CJ, Dobney K, Weyrich LS, Kaidonis J, Walker AW, Haak W, et al. Sequencing ancient calcified dental plaque shows changes in oral microbiota with dietary shifts of the Neolithic and Industrial revolutions. Nat Genet. 2013; 45: 450-5.
- Alberoni D, Gaggìa F, Baffoni L, Di Gioia D. Beneficial microorganisms for honey bees: problems and progresses. Appl Microbiol Biotechnol. 2016; 100: 9469-82.
- Araujo A, Reinhard KJ, Ferreira LF, Gardner SL. Parasites as probes for prehistoric human migrations? Trends Parasitol. 2008; 24: 112-5.
- Bakir B, Sari EK, Aydin BD, Yildiz SE. Immunohistochemical examination of effects of kefir, koumiss and commercial probiotic capsules on platelet derived growth factor-c and platelet derived growth factor receptor-alpha expression in mouse liver and kidney. Biotech Histochem. 2015; 90: 190-6.
- Bersaglieri T, Sabeti PC, Patterson N, Vanderploeg T, Schaffner SF, Drake JA, et al. Genetic signatures of strong recent positive selection at the lactase gene. Am J Hum Genet. 2004; 74: 1111-20.
- Botigué LR. Ancient European dog genomes reveal continuity since the Early Neolithic. Nature Communication 2017; 8:16082.
- Bourdichon F, Casaregola S, Farrokh C, Frisvad JC, Gerds ML, Hammes WP., et al. Food fermentations: microorganisms with technological beneficial use. Int J Food Microbiol. 2012; 154: 87-97.
- Bramanti B, Thomas MG, Haak W, Unterlaender M, Jores P, Tambets K, et al. Genetic discontinuity between local hunter-gatherers and central Europe’s first farmers. Science. 2009; 326: 137-40.
- Burger J, Kirchner M, Bramanti B, Haak W, Thomas MG. Absence of the lactase-persistence-associated allele in early Neolithic Europeans. Proc Natl Acad Sci USA. 2007; 104: 3736-41.
- Campbell-Platt G. Fermented Foods of the World: A Dictionary and Guide. Londres: Butterworths, 1987.
- Campbell-Platt G. Fermented foods – a world perspective. Food Res Int. 1994; 27: 253-7.
- Copley MS, Berstan R, Dudd SN, Docherty G, Mukherjee AJ, Straker V,et al. Direct chemical evidence for widespread dairying in prehistoric Britain. Proc Natl Acad Sci USA. 2003; 100: 1524-9.
- Corby-Harris V, Maes P, Anderson KE. The bacterial communities associated with honey bee (Apis mellifera) foragers. PLoS One. 2014; 9: e95056.
- Craig OE, Chapman J, Heron C, Willis LH, Bartosiewicz L, Taylor G, et al. Did the first farmers of central and eastern Europe produce dairy foods? Antiquity. 2005; 79: 882-94.
- Curry A. Archaeology: The milk revolution. Nature. 2013; 500: 20-2.
- De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci USA. 2010; 107: 14691-6.
- Dewan S, Tamang JP. Dominant lactic acid bacteria and their technological properties isolated from the Himalayan ethnic fermented milk products. Antonie Van Leeuwenhoek. 2007; 92: 343-52.
- Douglas GL, Klaenhammer TR. Genomic evolution of domesticated microorganisms. Annu Rev Food Sci Technol. 2010; 1: 397-414.
- Dunne J, Evershed RP, Salque M, Cramp L, Bruni S, Ryan K, et al. First dairying in green Saharan Africa in the fifth millennium BC. Nature. 2012; 486: 390-4.
- Edwards CJ, Bollongino R, Scheu A, Chamberlain A, Tresset A, Vigne JD, et al. Mitochondrial DNA analysis shows a Near Eastern Neolithic origin for domestic cattle and no indication of domestication of European aurochs. Proc Biol Sci. 2007; 274:1377-85.
- Ethier J, Bánffy E, Vuković J, Leshtakov K, Bacvarov K, Roffet-SalqueM, et al. Earliest expansion of animal husbandry beyond the Mediterranean zone in the sixth millennium BC. Sci Rep. 2017; 7: 7146.
- Evershed RP, Payne S, Sherratt AG, Copley MS, Coolidge J, Urem-Kotsu D, et al. Earliest date for milk use in the Near East and southeastern Europe linked to cattle herding. Nature. 2008; 455: 528-31.
- Franz CM, Huch M, Mathara JM, Abriouel H, Benomar N, Reid G, et al. African fermented foods and probiotics. Int J Food Microbiol. 2014; 190: 84-96.
- Gesudu Q, Zheng Y, Xi X, Hou QC, Xu H, Huang W, et al. Investigating bacterial population structure and dynamics in traditional koumiss from Inner Mongolia using single molecule real-time sequencing. J Dairy Sci. 2016; 99: 7852-63.
- Gibbons JG, Rinker DC. The genomics of microbial domestication in the fermented food environment. Curr Opin Genet Dev. 2015; 35:1-8.
- Gifford-Gonzalez D, Hanotte O. Domesticating animals in Africa: implications of genetic and archaeological findings. J World Prehist. 2011; 24: 1-23.
- Green JL, Holmes AJ, Westoby M, Oliver I, Briscoe D, Dangerfield M, et al. Spatial scaling of microbial eukaryote diversity. Nature 2004; 432: 747-50.
- Grieve J. An account of the method of making a wine, called by the tartars koumiss; with observations on its use in medicine. Transactions of the Royal Society of Edinburgh 1788; vol. I. Publicado al año siguiente en: Lond Med J. 1788; 10:197-210.
- Guillins MR, Paulsen IT. Microbiology of the Anthropocene. Anthropocene 2014; 5: 1-8.
- Horner-Devine MC, Lage M, Hughes JB, Bohannan BJ. A taxa–area relationship for bacteria. Nature 2004; 432: 750-3.
- Ismail MM, Hamad MF, Elraghy EM. Using goat’s milk, barley flour, honey, and probiotic to manufacture of functional dairy product. Probiotics Antimicrob Proteins. 2017; Aug 23. doi: 10.1007/s12602-017-9316-4.
- Itan Y, Powell A, Beaumont MA, Burger J, Thomas MG. The origins of lactase persistence in Europe. PLoS Comput Biol. 2009; 5: e1000491.
- Jagielski V. Koumiss in the treatment of phthisis. Br Med J. 1872; 1: 124-5.
- Jagielski VA. On the various preparations of koumiss, and their use in medicine. Br Med J. 1874; 1: 299-301.
- Jagielski V. The value of koumiss in the treatment of nausea, vomiting, and inability to retain other food on the stomach. Br Med J. 1877; 2: 919-21.
- Kelekna P. The horse in human history. Cambridge University Press; Cambridge: 2009.
- Kumari A, Angmo K, Monika, Bhalla TC. Probiotic attributes of indigenous Lactobacillus spp. isolated from traditional fermented foods and beverages of north-western Himalayas using in vitro screening and principal component analysis. J Food Sci Technol. 2016; 53: 2463-75.
- Kwong WK, Moran NA. Gut microbial communities of social bees. Nat Rev Microbiol. 2016; 14: 374-84.
- Librado P, Fages A, Gaunitz C, Leonardi M, Wagner S, Khan N, et al. The evolutionary origin and genetic makeup of domestic horses. Genetics. 2016; 204: 423-34.
- Litchman E. Invisible invaders: non-pathogenic invasive microbes in aquatic and terrestrial ecosystems. Ecol Lett. 2010; 13: 1560-72.
- Marshall F, Hildebrand E. Cattle before crops: the beginnings of food production in Africa. J World Prehist. 2002; 16: 99-143.
- Mayo B, Ammor MS, Delgado S, Alegría A. Fermented milk products. En: Fermented Foods and Beverages of the World, eds. JP Tamang, y K Kailasapathy, pp. 263-288. CRC Press, Taylor and Francis Group, Nueva York, 2010.
- McGovern PE, Zhang J, Tang J, Zhang Z, Hall GR, Moreau RA, et al. Fermented beverages of pre- and proto-historic China. Proc Natl Acad Sci USA. 2004; 101: 17593-8.
- McGovern P, Jalabadze M, Batiuk S, Callahan MP, Smith KE, Hall GR, et al. Early Neolithic wine of Georgia in the South Caucasus. Proc Natl Acad Sci USA. 2017; 114: E10309-18.
- Michelet L, Dauga C. Molecular evidence of host influences on the evolution and spread of human tapeworms. Biol Rev. 2012; 87: 731-41.
- Nakazawa Y, Hosono A. Fermented milk in the orient. En: Functions of fermented milk: challenges for the health sciences, eds. Y Nakazawa, A Hosono, pp. 61-78. Elsevier Applied Science, Londres, 1992.
- Outram AK, Stear NA, Bendrey R, Olsen S, Kasparov A, Zaibert V, et al. The earliest horse harnessing and milking. Science 2009; 323:1332-1335.
- Ozen M, Dinleyici EC. The history of probiotics: the untold story. Benef Microbes. 2015; 6: 159-65.
- Pan DD, Zeng XQ, Yan YT. Characterisation of Lactobacillus fermentum SM-7 isolated from koumiss, a potential probiotic bacterium with cholesterol-lowering effects. J Sci Food Agric. 2011; 91: 512-8.
- Pringle A, Bever JD, Gardes M, Parrent JL, Rillig MC, Klironomos JN. Mycorrhizal symbioses and plant invasions. Ann Rev Ecol Evol Syst. 2009; 40: 699-715.
- Rodríguez-Echeverría S. Rhizobial hitchhikers from Down Under: invasional meltdown in a plant–bacteria mutualism? J Biogeogr. 2010; 37: 1611-22.
- Roffet-Salque M, Regert M, Evershed RP, Outram AK, Cramp LJ, Decavallas O, et al. Widespread exploitation of the honeybee by early Neolithic farmers. Nature. 2015; 527: 226-30.
- Rong J, Zheng H, Liu M, Hu X, Wang T, Zhang X, et al. Probiotic and anti-inflammatory attributes of an isolate Lactobacillus helveticus NS8 from Mongolian fermented koumiss. BMC Microbiol. 2015; 15: 196.
- Salque M, Bogucki PI, Pyzel J, Sobkowiak-Tabaka I, Grygiel R, et al. Earliest evidence for cheese making in the sixth millennium BC in northern Europe. Nature. 2013; 493: 522-5.
- Shi T, Nishiyama K, Nakamata K, Aryantini NP, Mikumo D, Oda Y, et al. Isolation of potential probiotic Lactobacillus rhamnosus strains from traditional fermented mare milk produced in Sumbawa Island of Indonesia. Biosci Biotechnol Biochem. 2012; 76: 1897-903.
- Sicard D, Legras JL. Bread, beer and wine: yeast domestication in the Saccharomyces sensu stricto complex. C R Biol. 2011; 334: 229-36.
- Smith KF, Guégan J-F. Changing geographic distributions of human pathogens. Ann Rev Ecol Evol Syst. 2010; 41: 231-50.
- Steinkraus KH. Classification of fermented foods: worldwide review of household fermentation techniques. Food Control. 1997; 8: 331-17.
- Szécsényi-Nagy A, Roth C, Brandt G, Rihuete-Herrada C, Tejedor-Rodríguez C, Held P, et al. The maternal genetic make-up of the Iberian Peninsula between the Neolithic and the Early Bronze Age. Sci Rep. 2017; 7:15644.
- Tamang JP. Diversity of fermented foods. En: Fermented Foods and Beverages of the World, eds. JP Tamang, K Kailasapathy, pp. 41-84. CRC Press, Taylor and Francis Group, Nueva York, 2010a.
- Tamang JP. Diversity of fermented beverages. En: Fermented Foods and Beverages of the World, eds. JP Tamang, K Kailasapathy, pp. 85-125. CRC Press, Taylor and Francis Group, Nueva York, 2010a.
- Tamang JP, Tamang B, Schillinger U, Guigas C, Holzapfel WH. Functional properties of lactic acid bacteria isolated from ethnic fermented vegetables of the Himalayas. Int J Food Microbiol. 2009; 135: 28-33.
- Tamang JP, Watanabe K, Holzapfel WH. Diversity of microorganisms in globalfermented foods and beverages. Front Microbiol. 2016; 7: 377.
- Tamang JP, Shin DH, Jung SJ, Chae SW. Functional properties of microorganisms in fermented foods. Front Microbiol. 2016; 7: 578.
- Toussaint-Samat M. A History of Food. Wiley-Blackwell, Oxford, 2008
- Tsenina VS, Frolov VM. Use of koumiss made from cow’s milk in liver diseases. Meditsinskaia Sestra. 1980; 39: 29-30.
- Wang J, Liu L, Ball T, Yu L, Li Y, Xing F. Revealing a 5,000-y-old beer recipe in China. Proc Natl Acad Sci USA. 2016; 113: 6444-8.
- Walter J, Ley R. The human gut microbiome: ecology and recent evolutionary changes. Ann Rev Microbiol. 2011; 65: 411-29.
- Wilkinson DM. Have we underestimated the importance of humans in the biogeography of free-living terrestrial microorganisms? J Biogeogr. 2010; 37: 393-7.
- Wu R, Wang L, Wang J, Li H, Menghe B, Wu J, et al. Isolation and preliminary probiotic selection of lactobacilli from koumiss in Inner Mongolia. J Basic Microbiol. 2009; 49: 318-26.
- Wu R, Wang W, Yu D, Zhang W, Li Y, Sun Z, et al. Proteomics analysis of Lactobacillus casei Zhang, a new probiotic bacterium isolated from traditional home-made koumiss in Inner Mongolia of China. Mol Cell Proteomics. 2009; 8: 2321-38.
- Yao G, Yu J, Hou Q, Hui W, Liu W, Kwok LY, et al. A perspective study of koumiss microbiome by metagenomics analysis based on single-cell amplification technique. Front Microbiol. 2017; 8: 165.
- Yatsunenko T, Rey FE, Manary MJ, Trehan I, Domínguez-Bello MG, Contreras M, et al. Human gut microbiome viewed across age and geography. Nature. 2012; 486: 222-7.
- Zeder MA, Emshwiller E., Smith BD, Bradley DG. Documenting domestication: the intersection of genetics and archaeology. Trends Genet. 2006; 22: 139-55.
- Zhang W, Yu D, Sun Z, Wu R, Chen X, Chen W, et al. Complete genome sequence of Lactobacillus casei Zhang, a new probiotic strain isolated from traditional homemade koumiss in Inner Mongolia, China J Bacteriol. 2010; 192: 5268-9.
- Zhang J, Wang L, Guo Z, Sun Z, Gesudu Q, Kwok L, et al. 454 pyrosequencing reveals changes in the faecal microbiota of adults consuming Lactobacillus casei Zhang. FEMS Microbiol Ecol. 2014; 88: 612-22.
- Zhang J, Guo Z, Lim AAQ, Zheng Y, Koh EY, Ho D, et al. Mongolians core gut microbiota and its correlation with seasonal dietary changes. Sci Rep. 2014; 16: 5001.
- Zhao ZW, Pan DD, Wu Z, Sun YY, Guo YX, Zeng XQ. Antialcoholic liver activity of whey fermented by Lactobacillus casei isolated from koumiss. J Dairy Sci. 2014; 97: 4062-71.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}